DEZOKSİRİBONUKLEİN TURŞULARI (DNT) – canlı orqanizmdə genetik məlumatı saxlayan və ötürən yüksək molekullu təbii üzvi birləşmə. Ali orqanizmlərdə, bakteriya və bir çox viruslarda genetik məlumatın daşıyıcısı 2′dezoksiri bonukleotidlərdən ibarət olan biopolimerdir. DNT bir, yaxud ikizəncirli formada ola bilər və 2′dezoksiribonukleotidlər 3′, 5′fosfodiefir rabitələrlə birləşmişdir. İkizəncirli DNTdə iki komplementar zəncir ardıcıllığın əks istiqamətində (antiparalel orienta siya) birbirinin ətrafında burulmaqla sola və ya sağaburulan spiral əmələ gətirir. İki zəncir, azotlu əsaslar arasında hidrogen rabitələrinin spesifik yaranması, həmçinin stekinq qarşılıqlı təsir yolu ilə bir yerdə tutulur. Birzəncirli, yaxud ikizəncirli DNT həlqəvi molekul şəklində (mitoxondri, xloroplast, bakteriya) ola bilər. Bu hаlqа burularaq daha çox burulmuş spiral yarada bilir. Prokariot orqanizmlərdə DNT membranla birləşir və plazmid, yaxud episom formasında olur. Eukariot DNT-nin 95%-ə qədəri nüvədə yerləşir və spesifik zülallarla birləşmişdir. Mitoxondri DNT-si bütün hüceyrə DNT-sinin 1–2%-ni, xloroplast (plastid) DNT-sinin isə 5%-ə qədərini təşkil edir. Genetik məlumatın ana hüceyrədən qız hüceyrəyə köçürülməsi replikasiya nəticəsində baş verir. Növün bütün əlamətləri və fərdin xüsusiyyətləri haqqında genetik məlumatlar DNT-nin nukleotid ardıcıllığında
kodlaşmışdır. Bütün əsas genetik proseslərreplikasiya, transkripsiya, reparasiya və rekombinasiya DNT molekulunun fəaliyyəti ilə əlaqədardır.
İlk dəfə 1868 ildə İ.F. Mişer tərəfindən DNT zülallarla kompleks (dezoksiribonuk leoproteidlər) şəklində irin hüceyrələrinin nüvəsində və balıq spermalarında (1870) kəşf edilmişdir. 1889 ildə R. Altman nuklein turşularını zulallardan təmizləyərək, “nuklein”terminini biokimyaya gətirmişdir. Nuklein turşularının genetik rolu haqqında müasir təsəvvürlər ingilis mikrobioloqu F. Qriffits tərəfindən 1928 ildə Diplococcus pneumoniae bakteriyası ilə apardığı təcrübələrdən sonra formalaşmağa başlamışdır. Uzun müddət hesab edilirdi ki, DNT yalnız heyvan hüceyrələrində mövcuddur, lakin 1930 illərin ortalarında sübut edildi ki, (A.N. Belozerski) DNT hər bir canlı hüceyrənin əvəzolunmaz komponentidir. 1944 ildə Аmerika mikrobioloqu O.T. Eyveri əməkdaşları ilə göstərmişdir ki, irsi bioloji əlamət hüceyrənin bütün digər komponentlərindən əsaslı təmizlənmiş DNT-nin köməyilə bir hüceyrədən digərinə ötürülə bilər. Beləliklə, DNTnin bioloji funksiyası – onun irsiyyət materialı, yaxud genetik məlumatın daşıyıcısı olması müəyyənləşdirildi.
19 əsrin sonu – 20 əsrin əvvəllərində DNTnin monomer tərkibi – dezoksiriboza, fosfat turşusu və 4 azotlu əsasdan [purin əsasları – quanin (Q) və adenin (A); pirimidin əsasları – sitozin (S) və timin (T)] birinin qalıqları olan dezoksiribonukleotidlərdən ibarət polimer molekul olduğu müəyyən edildi. Eyni vaxtda E. Çarqaff əməkdaşları ilə DNT-nin nukleotid tərkibi nin əsas qanunauyğunluqlarını (Çarqaff qaydası) müəyyənləşdirdi ki, onlardan ən mühümü hər bir DNT molekulunda adenin və timin (A=T), həmçinin quanin və sitozin (G=S) qalıqlarının miqdarlarının bərabər olmasıdır.
Bu məlumatlara əsaslanaraq C.D. Uotson və F.H. Krik 1953 ildə R. Franklin və M. Uilkinsin laboratoriyalarında alınmış DNT kristallarının rentgenoqramlarının şifrəsini açmışlar. Onlar DNT molekulunun iki polinukleotid zəncirdən ibarət nizamlı (müntəzəm) quruluşu olan ikiqat spiral olmasını müəyyənləşdirdilər (şəkil 1). Spiralın diametri bütün uz. boyu sabitdir və təqr. 2 nm-ə bərabərdir. Spiralın burumunun uz. 3,4 nm-dir. Tək zəncirin hər burumunda təqr. 10 nukleotid qalığı yerləşir, yəni spiralın oxu boyunca nukleotidlərarası məsafə 0,34 nm-ə bərabərdir. Azotlu əsaslar DNT-nin ikiqat spiralında bir müstəvidə və praktiki olaraq spiralın əsas oxuna perpendi kulyar yerləşir. İki zəncirin qarşıqarşıya yerləşən azotlu əsasları hidrogen rabitələri ilə stabilləşmiş komplementar cütlər yaradır, beləliklə həmişə adenin yalnız timinlə, quanin isə sitozinlə cütləşir (Q–S cütü öz aralarında üç, A–T cütü isə yalnız iki hidrogen rabitəsi ilə birləşir, şəkil 2). DNT-nin ikiqat spiral quruluşunun stabilləşməsi üçün eyni bir zəncirə məxsus qonşu əsasların müstəviləri arasındakı qarşılıqlı təsir də mühüm əhəmiyyət kəsb edir.
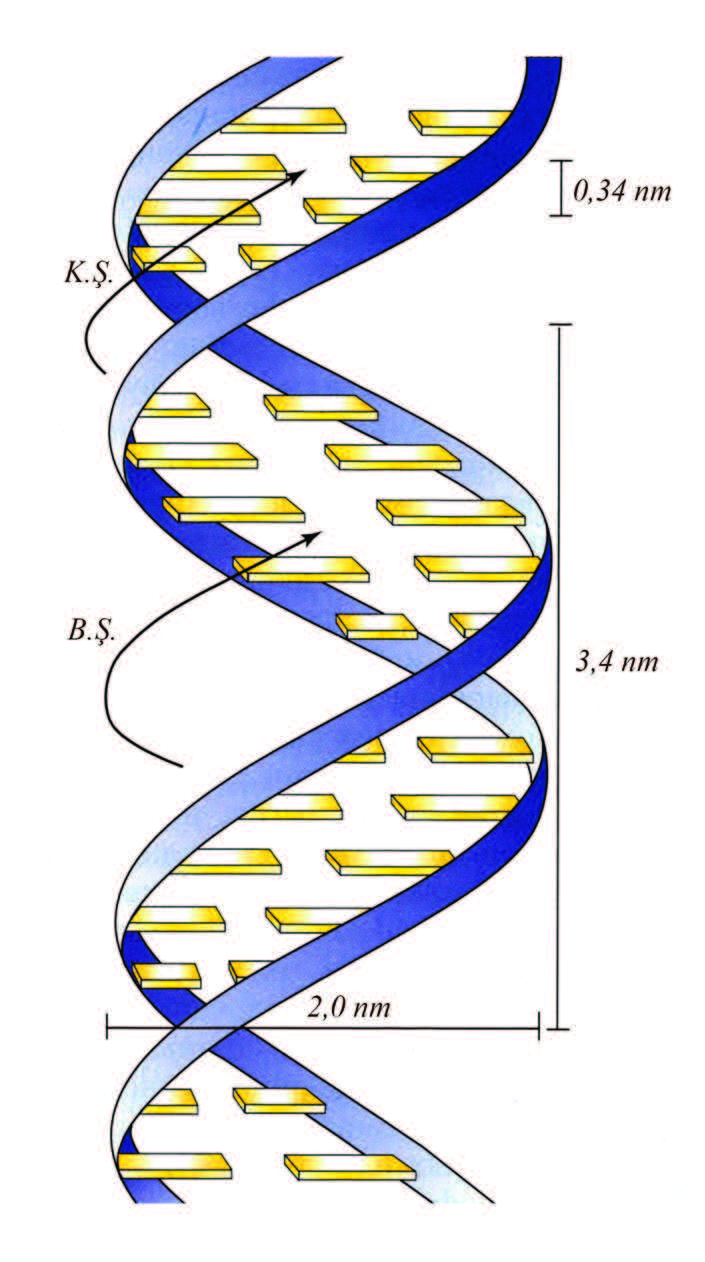 Шякил 1. ДНТ-нин икигат спиралынын Уотсон-Крик модели (К.Ш.– кичик шырым; Б.Ш.–бюйцк шырым).
Шякил 1. ДНТ-нин икигат спиралынын Уотсон-Крик модели (К.Ш.– кичик шырым; Б.Ш.–бюйцк шырым).
Uotson–Krikin ikiqat spiral modelindən DNT molekulunun (və yaxud, istənilən genetik materialın) özüözünü yaratması prinsipi (ikiləşmə, yaxud replikasiya) məlum olur: DNTnin iki komplementar zəncirini ayırıb, sonra matrisa üzərində onlara dəqiq komplementar olan yeni ikizəncirli iki molekul sintez olunur, bu iki ikizəncirli iki qız molekullar ana molekul ilə identik olur. Bu prinsipin kəşfi irsiyyət hadisəsinin molekulyar səviyyədə izah edilməsinə imkan yaratdı və molekulyar biologiyanın başlanğıcını qoymuş oldu. Nuklein turşuları əsaslarının komplementar cütləşməsi prinsipi hüceyrədə bütün genetik məlumatların ötürülməsi proseslərinin əsasını təşkil edir.
DNTnin ikiqat spiralında polinukleotid zəncirlərin şəkərfosfat skeleti xaricə yönəlmişdir, spiralın səthində isə iki şırım var: böyük şırımın eni 2,2 nm, kiçik şırımın eni isə 1,2 nm-dir. C. Uotson və F. Krikin təsvir etdiyi DNTnin ikiqat spiralı sağaburulmuş formadadır və onun polinukleotid zəncirləri isə antiparaleldir, yəni onlar əks istiqamətlərə elə yönəlmişlər ki, bir zəncirin 3′sonluğu digər zəncirinin 5′sonluğu qarşısında yerləşir və Bforması adlanır. DNTnin B fəsiləsinə CDNT və DDNT formaları da daxildir. Genin aktivliyinin tənzimlənmə sində DNT polimorfizmi mühüm rol oynaya bilər və Bformadan başqa DNTnın ikiqat spiralının digər konformasiyalarının invivo olması haqqında artıq yeni məlumatlar vardır.
DNTnin ikiqat spiralı əhəmiyyətli də rəcədə polimorfizmlə xarakterizə olunur və xarici şərait dəyişdikdə Uotson-Krikin B formasından fərqli fəza quruluşu (konformasiya) ala bilir. Belə ki, preparatda rütubət azaldıqda, yaxud məs., DNTnin sulu məhluluna etanol əlavə edildikdə Bforma A formaya keçir.
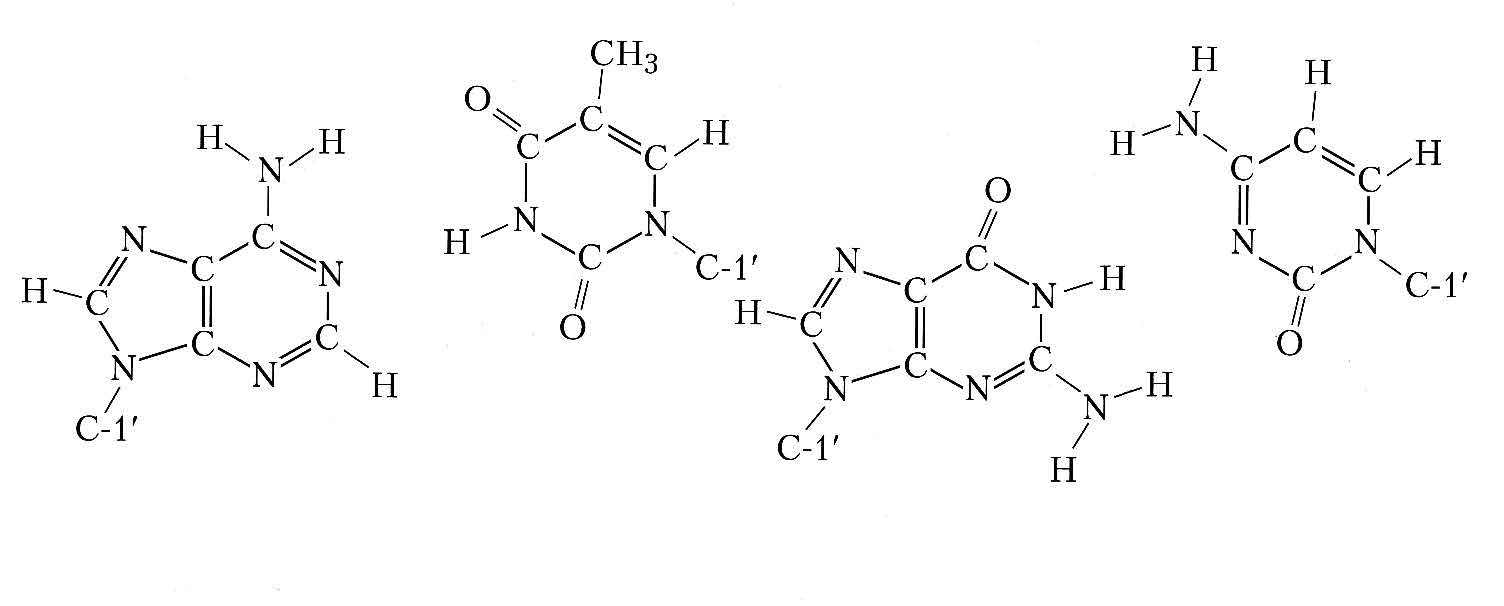
Шякил 2. Икиспираллы ДНТ молекулунда комплементар Уотсон-Крик ъцтц. Солда аденин-тимин ъцтц;саьда гуанин-ситозин ъцтц. Спесифик щидроэен рабитяляри иля бирляшмиш атомлар, щямчинин дезоксирибоза галыгларынын Ъ-1′ атомлары арасындакы мясафяляр верилмишдир
ADNT forması həmçinin Uotson-Krik nukleotid cütləri ilə stabilləşmiş sağaburulan ikiqat zəncirdir. BDNT-dən fərqli ola raq ADNT aşağı nisbi rütubət şəraitində yaranır. Biomolekulun kristallaşmasını stimullaşdıran reagentlər onun dehidratasiyasına səbəb olur və bir çox DNT molekulu kristal halında Aformaya keçir. Aformada antiparalel polinukleotid zəncirlər BDNT nisbətən az dərəcədə burulmuşlar. Spiralın bir burumuna 11–12 nukleotid cütü düşür. ADNT C3′-endo-, BDNT isə C2′-endo- konformasiyaya malikdir. Әlverişsiz şəraitdə bəzi bakteriyalar sporlara çevrilirlər və belə qeyri-aktiv vəziyyətdə uzun müddət qala bilirlər. Onların DNT-si spor zülalları ilə əhatə edilmiş və ADNT formasında olur. Bu halda DNT molekulu UB şüanın təsirinə 10 dəfə çox davamlı olur. Spor zülalları Bformadan Aformaya çevrilməkdə bakteriya DNT-sinə kömək edir və əlverişsiz şəraitdə DNT-ni mühafizə edir.
DNTnin ikiqat spirallarının mühüm xassələrindən biri onların yüksək aşkarlayıcı rentgenstruktur analiz ilə aşkar olunan mikroheterogenliyidir. Bu, nukleotid qalıqlarının konformasiyasındakı incə fərqlərlə şərtlənir ki, onların görünməsi zəncirdə nukleotidlərin yerləşmə ardıcıllığından asılıdır və xarakterik bükük və sınmaların əmələ gəlməsi ilə meydana çıxır. DNT molekulunun quruluşunun belə xüsusiyyətləri şübhəsiz ki, onun fəaliyyəti ilə əlaqədardır.
DNT molekulunda təkrarlanan nukleotid ardıcıllıqları (palindromlar) olan zaman cütlər yalnız qarşıqarşıya olan (əks) zəncirlərin əsasları arasında deyil, həm də bir zəncir çərçivəsində əsaslar arasında formalaşa bilir ki, bu da hidrogen rabitələri ilə birləşmiş özünəməxsus sancaq ilgəklərinin yaranmasına imkan yaradır.
DNT məhlulların qızdırılması, pH-ın dəyişməsi, ion quvvəsinin azalması, bir sıra üzvi maddələrin, digər birləşmələrin və faktorların təsiri ilə DNT-nin denaturasiyası – cüt əsaslar arasındakı hidrogen rabitələrinin qırılması və ikiqat spiralın müntəzəm quruluşunun dağılması hesabına zəncirlər birbirindən tam ayrılır. İkiqat spiralı stabilləşdirən molekuldaxili qarşılıqlı təsirlərin kooperativ xarakteri hesabına bu proses faza keçidini xatırladır və bu na görə DNT-nin əriməsi adlanır. İkiqat spiralın əmələ gəlməsi üçün optimal şərait də DNT-nin ayrı-ayrı komplementar zəncirləri əvvəlki ikiqat spiral quruluşu bərpa etməklə (DNT-nin renaturasiyası) reasso siasiya qabiliyyətinə malikdir. Termiki denaturasiya, yaxud ərimə 80 – 90°C-də baş verir, soyuyan zaman isə ikiqat DNT molekulu bərpa olunur. Bu xassə DNT, yaxud DNT və RNT molekullarının nukleotid ardıcıllıqlarında oxşarlıq dərəcəsini, onların təşkil xüsusiyyətlərini, o cümlədən təkrarların mövcudluğunu və sayını aşkar etməyə imkan verən nuklein turşularının molekulyar hibridləşmə metodunun əsasında durur.
Müxtəlif orqanizmlərin DNT molekulunda nukleotid qalıqlarının növbələşən ardıcıllığı (birinci quruluş) qəti olaraq fərdidir, DNTnin bir molekulunu digərindən və uyğun olaraq bir geni, yaxud tənzimləyici genetik elementi digərindən fərqləndirən mühüm xüsusiyyətidir. DNT molekullarının ölçüsü bir neçə min nukleotid cütündən (plazmid və bəzi viruslar) yüz minlərlə cütə qədər (ali orqanizmlərdə) dəyişir. Müxtəlif orqanizmlərdə DNTnin miqdarı da fərqlidir, onu əmələ gətirən nukleotidlərin sayına görə bakteriyalarda 5·106, ali bitki genomlarında isə 2·1011 nukleotid cütü təşkil edir (haploid genomuna görə hesablanmış). Bu nəhəng molekullar hüceyrə, yaxud viruslarda çox kompakt qablaşmışdır. Prokariot nukleotiddə belə qablaşma az miqdarda xüsusi zülallar və ribonuklein turşularının (RNT) hesabına təmin olunduğu ehtimal edilir. Universal histon və bəzi qeyrihiston zülallar dəstinin köməyilə xromosomun əsas komponentinin-xromatinin yaranmasına səbəb olan eukariot DNT-nin qablaş masının bir neçə səviyyəsi xarakterizə olunmuşdur. Məs., insanın ən böyük xromosomunun DNT-nin uz. 8 sm olduğu halda, mitoz zamanı spirallaşmış vəziyyətdə xromosomda onun uz. 5 mkm-dan çox deyildir.
Eukariotların somatik hüceyrələrinin nüvələrində DNT molekulu iki nüsxədə olur. Hər bir pro- və eukariot xromosomun da yalnız bir ikispirallı DNT molekulu var. Virusların müəyyən qisminin genomları həmçinin ikispirallı DNT ilə təmsil olunmuş və yalnız bəzi faqların genomu birzəncirli hаlqаvi, yaxud xətti DNT molekulundan ibarətdir.
Prokariot xromosomların, plazmidlərin, bir çox virusların, mitoxondrilərin və xloroplastların ikisaplı DNT molekulları həlqəvi qapanmışdır. Bu zaman əgər zəncir kovalentfasiləsizdirsə (yəni bütün fosfodiefir rabitələri qapanmışsa), onda ikiqat spiralın sapları bir neçə dəfə bir-biri ilə kəsişdikləri, yaxud birləşdikləri üçün tsiklik DNT yüksəkspirallaşmış formada ola bilər. Hüceyrədə superburumlar topoizomeraza fermentlərinin iştirakı ilə yaranır və dağılır. Tsiklik yüksəkspirallaşmış DNT onun xətti formasına nisbətən müəyyən enerji ehtiyatına malikdir və buna görə superburmaların yaranması DNT-nin fəaliyyəti üçün vacibdir (məs., replikasiya zamanı meydana çıxan topoloji çətinlikləri aradan götürməyə imkan verir). Bundan başqa, superburmaların olması hesabına onun makromolekulunda qeyriadi quruluşlar yarana bilər: xaçabənzər quruluşlar (palindromlarda), Zforma, üçsaplı sahələr, yaxud Hforma (homopurinhomopirimidin bloklarda) yarana bilər.
DNT-nin biosintezi (replikasiya) DNT polimeraza fermentlərin və köməkçi zülalların böyük bir qrupunun birgə iştirakı zamanı matrisa sintezi yolu ilə həyata keçirilir və hüceyrənin xüsusi tənzimləyici sistemlərinin nəzarəti altında gedir. DNT-nin istənilən sahəsi Polimeraza Zəncir Reaksiyası ilə in vitro amplifikasiya oluna bilər. Replikasiyanın in vivo gedişində, həmçinin onun sona çatmasından sonra az sayda müəyyən sitozin qalıqlarının 5 metilsitozin əmələ gəlməsilə nəticələnən metilləşməsi baş verir, bu DNT modifi kasiyasının spesifik prosesini təmsil edir və onun sonrakı fəaliyyəti ilə bilavasitə bağlıdır. DNT-nin metilləşməsi və metilsizləşməsi embrio və qametogenez proseslərində mühüm rol oynayır.
Orqanizmlərin həyat fəaliyyətində onların DNT-si xarici amillərin təsiri altında azotlu əsasların quruluşunun pozulması ilə müşayiət olunan müxtəlif zədələyici təsirlərə məruz qala bilər. Təkamülün gedişində hüceyrələr DNT-nin ilkin quruluşunun bərpasını təmin edən (reparasiya) müdafiə mexanizmlərini inkişaf etdirmişdir.
Hüceyrədə DNT spesifik fermentlər dezoksiribonukleazalarla parçalanır. Bunların arasında ən çox məlum olan hüceyrəni yad DNT-dən qoruyan və gen mühəndisli yində geniş istifadə olunan restriksiya endo nukleazalarıdır.
1970 illərin əvvəllərində F. Senger və başqaları tərəfindən DNT molekulların nukleotid ardıcıllıqlarını təyin edən səmərəli metodlar hazırlanmışdır. 20 əsrin sonunda bu metodların əsasında DNT-nin sekvens edilməsi üçün güclü avtomatlaşdırılmış texnologiya yaradılmış və onun vasitəsilə bir çox virusların, mitoxondrilərin, xloroplastların, bakteriyaların, bitki və heyvanların tam genomlarının DNT-nin birinci quruluşu təyin edilmişdir. 2004 ilə qədər insanın tam genomunun nukleotid ardıcıllığının təyin edilməsi başa çatdırıldı. Bu işlər bioinformatikanın inkişafına təkan vermiş və molekulyar genetikanın yeni bölməsi olan genomikanın əsasını qoymuşdur.
DNTnin nukleotid ardıcıllığı haqqında məlumatlar rekombinant DNT molekullarının, yəni əvvəlcədən müəyyən olunmuş xassələrə malik molekulun yaradılması zamanı, müxtəlif orqanizmlərin DNT-nin quruluş elementləri daxil edilmiş molekulların (bax Gen mühəndisliyi), həmçinin yeni zülalların konstruksiyası zamanı geniş istifadə olunur. DNT-nin birinci quruluşu haqqında biliklər irsi və onkoloji xəstəliklərin analizi, şəxsiyyətin identifikasiyası, müəyyən genlərinin amplifikasiyası və ayrılması, tənzimləyici elementlərin və DNT-nin funksional mühüm sahələrinin analizi zamanı vacibdir.
Əd.: W a t s o n J.D., C r i c k F.H.C. Molecular structure of nucleic acid. Nature, 1953, vol. 171; У о т с о н Дж. Молекулярная биология гена. М., 1967; Х а г г и с Дж., М и х и Д., М ю и р А., Р о б е р т с К., У о к е р П. Введение в молекулярную биоло гию. М., 1967; А г о л В.И., Б о г д а н о в А.А., Г в о з д е в В.А., Г р а г е р о в А.И., К о л ч и н с к и й А.М., М и р з а б е к о в А.Д., Никифоров В.Г. Молекулярная биология. Структура и био синтез нуклеиновых кислот. М., 1990; С. М ю л л е р. Нуклеиновые кислоты, От А до Я. М., 2013.














